단백질비생성성 아미노산
단백질비생성성 아미노산(영어: non-proteinogenic amino acid)은 생화학에서 단백질의 생합성을 위해 생명체의 게놈(유전체)에 자연적으로 암호화되어 있는 22가지의 단백질생성성 아미노산(진핵생물의 경우 21가지[주 1])이 아닌 아미노산들을 지칭한다. 그러나 140가지 이상의 단백질비생성성 아미노산이 단백질에서 자연적으로 생성되며, 수천 가지 이상의 단백질비생성성 아미노산이 자연에서 생성되거나 실험실에서 합성될 수 있다.[1] 다음과 같은 역할로 인해 많은 단백질비생성성 아미노산들은 중요하다.
- 생합성에서의 대사 중간생성물,
- 단백질의 번역 후 변형에서,
- 생리학적 역할에서 (예: 세균의 세포벽의 구성 요소, 신경전달물질 및 독소),
- 천연 또는 인공 약리학적 화합물,
- 운석에 존재하거나 생물 발생 이전에 대한 실험 (예: 밀러와 유리의 실험)에 사용된다.

존재의 부정에 대한 정의 편집

기술적으로 아미노기(–NH2)와 카복실기(–COOH)를 가지고 있는 모든 유기 화합물은 아미노산이다. 단백질생성성 아미노산은 아미노기, 카복실기, 곁사슬, α-수소의 L-입체형태를 갖는 중심 탄소 원자(α-탄소 또는 2-탄소)를 가지고 있는 부류의 작은 부분 집합이다. 비카이랄성인 글리신과 아미노기가 2차 아민인 프롤린은 결과적으로 이민은 아니지만 전통적인 이유로 인해 종종 이미노산으로 불린다.
번역 중에 단백질에 통합되는 20가지의 표준 아미노산들은 유전 부호로 암호화되어 있다. 그러나 셀레노시스테인과 피롤리신이라는 두 가지 추가적인 단백질생성성 아미노산이 있다. 이러한 비표준 아미노산은 전용 코돈이 없지만 특정 서열이 존재할 때 종결 코돈 대신에 첨가되며, 셀레노시스테인의 경우 UGA 코돈과 SECIS 요소,[2] 피롤리신의 경우 UAG 코돈과 PYLIS 하류 서열을 필요로 한다.[3] 다른 모든 아미노산들은 단백질비생성성 아미노산이다.
-
 셀레노시스테인의 구조. 이 β-탄소에 셀레놀기를 포함하고 있다.
셀레노시스테인의 구조. 이 β-탄소에 셀레놀기를 포함하고 있다. -
 피롤리신의 구조. 이 아미노산은 리신의 ε-아미노기에 카복실화된 피롤린 고리가 결합하여 형성된다.
피롤리신의 구조. 이 아미노산은 리신의 ε-아미노기에 카복실화된 피롤린 고리가 결합하여 형성된다.

다음과 같이 다양한 아미노산의 부류들이 있다.[4]
- 20가지의 표준 아미노산
- 22가지의 단백질생성성 아미노산
- 고농도에서 무생물적으로 생성되는 80가지 이상의 아미노산
- 약 900가지는 자연적인 경로에 의해 생성된다.
- 118가지 이상의 인공적인 아미노산이 단백질에 삽입되었다.
이들 부류들은 서로 겹치는 부분이 있을 수 있지만 동일하지는 않다. 22가지의 모든 단백질생성성 아미노산은 생물에 의해 생합성되며 전부는 아니지만 일부는 비생물적(생물 발생 이전에 관한 실험 및 운석에서 발견됨)이기도 하다. 노르류신과 같은 일부 천연 아미노산은 단백질 합성 과정의 오류로 인해 번역시 단백질로 잘못 삽입되기도 한다. 오르니틴과 같은 많은 아미노산들은 생합성으로 생성되는 대사 중간생성물이지만 번역시 단백질에 삽입되지는 않는다. 단백질에서 아미노산 잔기들은 번역 후 변형을 통해 단백질생성성 아미노산에서 단백질비생성성 아미노산으로 변형된다. 다른 아미노산들은 비생물적 혼합물(예: α-메틸노르발린)에서만 발견된다. 30가지 이상의 비천연 아미노산들이 유전공학적 방법에 의해 번역시 단백질로 삽입되지만 생합성되지는 않았다.[4]
명명법 편집
유기 분자의 다양한 탄소를 구별하기 위한 IUPAC 번호 매기기 체계 외에도 카복실기를 구성하는 탄소의 다음 탄소부터 각 탄소에 순차적으로 그리스 문자를 표시하여 구분할 수도 있다. 여기서 α-탄소는 α-아미노산에서 아미노기, 카복실기, 곁사슬, 수소 원자와 결합하고 있는 중심 카이랄 탄소이다. 그리스 문자로 나타낼 시에 카복실기의 탄소는 고려하지 않는다.[5] 따라서 많은 단백질비생성성 아미노산들의 IUPAC 이름은 "2-아미노-"로 시작하여 "-산"으로 끝난다.
천연 비-L-α-아미노산 편집
대부분의 천연 아미노산은 L-입체형태의 α-아미노산이지만 일부 예외가 존재한다.
비-α-아미노산 편집

일부 비-α-아미노산이 생물에 존재한다. 이러한 구조에서 아미노기는 아미노산 분자의 카복실기로부터 더 먼 곳에 위치한다. 따라서 β-아미노산은 카복실기의 탄소로부터 두 번째 탄소에 아미노기가 결합되어 있고, γ-아미노산은 세 번째 탄소에 아미노기가 결합되어 있다. 예로는 β-알라닌, γ-아미노뷰티르산(GABA), δ-아미노레불린산이 있다.
-

-
 γ-아미노뷰티르산(GABA): 동물의 신경전달물질이다.
γ-아미노뷰티르산(GABA): 동물의 신경전달물질이다. -

-




α-아미노산이 단백질에 사용되는 이유는 운석 및 생물 발생 이전과 관련된 실험에서의 빈도와 관련이 있다.[7] 2차 구조의 측면에서[7] β-아미노산의 유해한 특성에 대한 초기 추측은 잘못된 것으로 밝혀졌다.[8]
D-아미노산 편집
일부 아미노산에는 정상적인 리보솜에 의한 번역 및 전사에 사용할 수 없는 카이랄성이 반대인 화학 물질이 포함되어 있다. 대부분의 세균의 세포벽은 아미노당으로 구성된 중합체가 서로 짧은 올리고펩타이드로 가교된 펩티도글리칸으로 구성되어 있다. 올리고펩타이드는 비리보솜으로 합성되며 D-아미노산들, 일반적으로 D-알라닌과 D-글루탐산을 포함하는 여러 특성들을 가지고 있다. 추가적인 특징은 전자가 피리독살 인산(PLP) 결합 효소(alr 또는 상동유전자인 dadX에 의해 암호화 됨)에 의해 라세미화되는 반면 후자는 보조 인자에 대해 독립적인 효소(murI에 의해 암호화 됨)에 의해 라세미화된다는 것이다. 일부 변형이 존재하는 데, 테르모토가속(Thermotoga)의 종에는 D-리신이 존재하며, 특정 반코마이신 내성 세균에는 D-세린이 존재(vanT 유전자)한다.[9][10]
α-탄소에 수소가 없는 경우 편집
모든 단백질생성성 아미노산은 α-탄소에 적어도 하나의 수소를 가지고 있다. 글리신은 두 개의 수소를 가지고 있고, 다른 단백질생성성 아미노산은 한 개의 수소와 한 개의 곁사슬을 가지고 있다. 수소를 메틸기와 같은 더 큰 치환기로 치환하면 단백질 골격이 왜곡된다.[7]
일부 균류에서 α-아미노아이소뷰티르산은 펩타이드의 전구체로 생성되며, 그 중 일부는 항생제의 특성을 나타낸다.[11] 이 화합물은 알라닌과 유사하지만 α-탄소에 수소 대신에 메틸기를 가지고 있다. 따라서 α-아미노아이소뷰티르산은 카이랄성이 아니다. α-수소가 없는 알라닌과 유사한 또 다른 화합물로는 메틸렌 곁사슬을 가지고 있는 디하이드로알라닌이 있다. 이것은 자연적으로 생성되는 여러 디하이드로아미노산들 중 하나이다.
-
 알라닌의 구조
알라닌의 구조 -
 아미노아이소뷰티르산의 구조
아미노아이소뷰티르산의 구조 -
 디하이드로알라닌의 구조
디하이드로알라닌의 구조



트윈 아미노산 입체중심 편집
L-α-아미노산의 하위 집합은 두 말단 중 어느 쪽이 α-탄소인지 모호하다. 단백질에서 시스테인 잔기는 다른 시스테인 잔기와 이황화 결합을 형성하여 단백질을 가교할 수 있다. 두 개의 가교된 시스테인은 시스틴을 형성한다. 시스테인과 메티오닌은 일반적으로 직접적인 황산화에 의해 생성되지만 일부 종에서는 활성화된 호모세린 또는 세린이 시스테인 또는 호모시스테인과 융합하여 시스타티오닌을 형성하는 황전환 경로에 의해 생성될 수 있다. 유사한 화합물로는 싸이오에스터 결합을 통해 연결된 두 개의 알라닌 분자로 볼 수 있는 란티오닌이 있으며, 이것은 다양한 생물들에서 발견된다. 유사하게 젱콜콩의 식물 독소인 젠콜산은 메틸렌기에 의해 연결된 두 개의 시스테인으로 구성된다. 다이아미노피멜산은 펩티도글리칸에서 가교로 사용되며, 리신의 전구체로 (탈카복실화를 통해) 사용된다.
-
 시스틴의 구조
시스틴의 구조 -
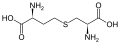 시스타티오닌의 구조
시스타티오닌의 구조 -
 란티오닌의 구조
란티오닌의 구조 -
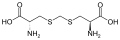 젠콜산의 구조
젠콜산의 구조 -
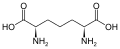 다이아미노피멜산의 구조
다이아미노피멜산의 구조

생물 발생 이전의 아미노산 및 대체생화학 편집
운석과 생물 발생 이전의 실험(예: 밀러와 유리의 실험)에서는 20가지의 표준 아미노산보다 더 다양한 아미노산들이 발견되며, 그 중 일부는 표준 아미노산보다 더 높은 농도에서 발견된다. 아미노산에 기반한 생명체가 우주의 다른 곳에서 평행적으로 발생하였다면 아미노산의 75% 이상은 공유되지 않을 것이라고 추측되었다.[7] 가장 주목할만한 이상은 아미노뷰티르산의 부족이다.
| 분자명 | 방전 | 머치슨 운석 |
|---|---|---|
| 글리신 | 100 | 100 |
| 알라닌 | 180 | 36 |
| α-아미노-n-뷰티르산 | 61 | 19 |
| 노르발린 | 14 | 14 |
| 발린 | 4.4 | |
| 노르류신 | 1.4 | |
| 류신 | 2.6 | |
| 아이소류신 | 1.1 | |
| 알로아이소류신 | 1.2 | |
| t-류신 | < 0.005 | |
| α-아미노-n-헵탄산 | 0.3 | |
| 프롤린 | 0.3 | 22 |
| 피페콜산 | 0.01 | 11 |
| α,β-다이아미노프로피온산 | 1.5 | |
| α,γ-다이아미노뷰티르산 | 7.6 | |
| 오르니틴 | < 0.01 | |
| 리신 | < 0.01 | |
| 아스파르트산 | 7.7 | 13 |
| 글루탐산 | 1.7 | 20 |
| 세린 | 1.1 | |
| 트레오닌 | 0.2 | |
| 알로트레오닌 | 0.2 | |
| 메티오닌 | 0.1 | |
| 호모시스테인 | 0.5 | |
| 호모세린 | 0.5 | |
| β-알라닌 | 4.3 | 10 |
| β-아미노-n-뷰티르산 | 0.1 | 5 |
| β-아미노아이소뷰티르산 | 0.5 | 7 |
| γ-아미노뷰티르산 | 0.5 | 7 |
| α-아미노아이소뷰티르산 | 7 | 33 |
| 아이소발린 | 1 | 11 |
| 사르코신 | 12.5 | 7 |
| N-에틸글리신 | 6.8 | 6 |
| N-프로필글리신 | 0.5 | |
| N-아이소프로필글리신 | 0.5 | |
| N-메틸알라닌 | 3.4 | 3 |
| N-에틸알라닌 | < 0.05 | |
| N-메틸-β-알라닌 | 1.0 | |
| N-에틸-β-알라닌 | < 0.05 | |
| 아이소세린 | 1.2 | |
| α-hydroxy-γ-aminobutyric acid | 17 |
직선형 곁사슬 편집
유전 부호는 동결 사건으로 설명되었으며 직선형 사슬이 있는 표준 아미노산(알라닌)이 하나만 있는 이유는 단순히 발린, 류신, 아이소류신이 중복되기 때문일 수 있다.[7] 그러나 직선형 아미노산은 훨씬 더 안정적인 α-나선을 형성하는 것으로 보고되었다.[12]
-
 글리신의 구조 (곁사슬이 수소 원자)
글리신의 구조 (곁사슬이 수소 원자) -
알라닌의 구조 (곁사슬이 메틸기)
-
 호모알라닌 또는 α-아미노뷰티르산의 구조 (곁사슬이 에틸기)
호모알라닌 또는 α-아미노뷰티르산의 구조 (곁사슬이 에틸기) -
 노르발린의 구조 (곁사슬이 n-프로필기)
노르발린의 구조 (곁사슬이 n-프로필기) -
 노르류신의 구조 (곁사슬이 n-뷰틸기)
노르류신의 구조 (곁사슬이 n-뷰틸기) -
 호모노르류신의 구조 (곁사슬이 n-펜틸기)
호모노르류신의 구조 (곁사슬이 n-펜틸기)




16족 원소 (칼코젠) 편집
세린, 호모세린, O-메틸호모세린 및 O-에틸호모세린은 하이드록시메틸기, 하이드록시에틸기, O-메틸하이드록시메틸기 및 O-메틸하이드록시에틸기가 있는 곁사슬을 가지고 있으며, 시스테인, 호모시스테인, 메티오닌 및 에티오닌은 싸이올 등가물을 가지고 있다. 셀레놀 등가물은 셀레노시스테인, 셀레노호모시스테인, 셀레노메티오닌, 셀레노에티오닌이다. 다음과 같은 16족 원소를 가지고 있는 아미노산들도 자연에서 발견된다. 아스페르길루스 푸미가투스(Aspergillus fumigatus) 아스페르길루스 테레우스(Aspergillus terreus) 및 페니실리움 크리소게눔(Penicillium chrysogenum)과 같은 여러 생물종들은 황이 없는 상태에서 텔루로시스테인 및 텔루로메티오닌을 생성하고 단백질로 통합될 수 있다.[13]
확장된 유전 부호 편집
역할 편집
세포 특히 독립영양생물에서 여러 단백질비생성성 아미노산들이 대사 중간생성물로 발견된다. 그러나 피리독살 인산(PLP) 결합 효소의 촉매적 유연성에서 불구하고 많은 아미노산들이 케토산(예: 4-메틸-2-옥소펜탄산에서 류신으로)으로 합성되고 마지막 단계에서 아미노화되어 단백질비생성성 아미노산의 대사 중간생성물의 수를 상당히 낮게 유지한다.
오르니틴과 시트룰린은 아미노산 이화작용의 일부인 요소 회로에서 생성된다(아래 참조).[14]
1차 대사에 더하여, 몇몇 단백질비생성성 아미노산은 작은 화합물 또는 비리보솜 펩타이드(일부 독소와 같은)를 만들기 위한 2차 대사의 전구체 또는 최종 생성물이다.
번역 후 단백질로 통합 편집
유전 부호에 의해 단백질생성성 아미노산으로 암호화되지 않음에도 불구하고 일부 비표준 아미노산들이 단백질에서 발견된다. 이들은 표적 단백질에 존재하는 표준 아미노산의 곁사슬의 번역 후 변형에 의해 형성된다. 이러한 변형은 보통 단백질의 기능이나 조절에 필수적이다. 예를 들어 γ-카복시글루탐산에서 글루탐산의 카복실화는 칼슘 양이온과 더 잘 결합하게 하고,[15] 하이드록시프롤린에서 프롤린의 하이드록실화는 결합 조직을 유지하는 데 중요하다.[16] 또 다른 예는 리신 잔기의 변형을 통한 번역 개시인자인 EIF5A에서 하이퓨신의 형성이다.[17] 이러한 변형은 또한 단백질의 국소화를 결정할 수 있으며, 예를 들어 긴 소수성 작용기를 첨가하면 단백질을 인지질 막에 결합시킬 수 있다.[18]
-
 카복시글루탐산의 구조. 글루탐산은 1개의 γ-카복실기를 가지고 있는 반면, 카복시글루탐산은 2개의 γ-카복실기를 가지고 있다.
카복시글루탐산의 구조. 글루탐산은 1개의 γ-카복실기를 가지고 있는 반면, 카복시글루탐산은 2개의 γ-카복실기를 가지고 있다. -
 하이드록시프롤린의 구조. 이 이미노산은 고리의 4번 탄소의 하이드록실기로 인해 프롤린과 다르다.
하이드록시프롤린의 구조. 이 이미노산은 고리의 4번 탄소의 하이드록실기로 인해 프롤린과 다르다. -
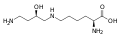
-
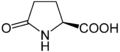 피로글루탐산의 구조
피로글루탐산의 구조


-Pyroglutamic_acid_Structural_Formulae.png)
독성 유사체 편집
싸이알리신과 같은 몇몇 단백질비생성성 아미노산은 단백질생성성 아미노산의 특정 특성을 모방하는 능력으로 인해 독성이 있다. 퀴스쿠알산, 카나바닌, 아제티딘-2-카복실산을 비롯한 일부 단백질비생성성 아미노산은 신경전달물질(즉, 단백질 생합성에 사용되지 않음)로 사용되는 아미노산을 모방하여 신경독성을 나타낸다.[21] 세팔로스포린 C는 세팔로스포린 부분으로 아마이드화된 α-아미노아디프산(호모글루탐산) 골격을 가지고 있다.[22] 페니실라민은 작용 방식이 알려지지 않은 치료용 아미노산이다.
-
 싸이알리신의 구조
싸이알리신의 구조 -
 퀴스쿠알산의 구조
퀴스쿠알산의 구조 -
 카나바닌의 구조
카나바닌의 구조 -
 아제티딘-2-카복실산의 구조
아제티딘-2-카복실산의 구조 -
 세팔로스포린 C의 구조
세팔로스포린 C의 구조 -
 페니실라민의 구조
페니실라민의 구조

-Azetidine-2-carboxylate.png)

자연적으로 생성되는 사이아노톡신에는 단백질비생성성 아미노산들도 포함될 수 있다. 예를 들어 마이크로시스틴과 노둘라린은 둘 다 β-아미노산인 ADDA로부터 유도된다.
아미노산이 아닌 것들 편집
타우린은 아미노설폰산이며 아미노카복실산이 아니지만 때때로 특정 생물체(고양이와 같은)에서 영양요구성을 억제하는 데 필요한 양이 비타민(보조 인자 영양요구성)보다 "필수 아미노산"(아미노산 영양요구성)에 더 가까운 것으로 간주된다.
삼투물질인 사르코신 및 트라이메틸글리신은 아미노산으로부터 유도되지만 각각 2차 아민 및 4차 아민을 가지고 있다.
같이 보기 편집
주해 편집
각주 편집
- ↑ Ambrogelly, A.; Palioura, S.; Söll, D. (2007). “Natural expansion of the genetic code”. 《Nature Chemical Biology》 3 (1): 29–35. doi:10.1038/nchembio847. PMID 17173027.
- ↑ Böck, A.; Forchhammer, K.; Heider, J.; Baron, C. (1991). “Selenoprotein synthesis: An expansion of the genetic code”. 《Trends in Biochemical Sciences》 16 (12): 463–467. doi:10.1016/0968-0004(91)90180-4. PMID 1838215.
- ↑ Théobald-Dietrich, A.; Giegé, R.; Rudinger-Thirion, J. L. (2005). “Evidence for the existence in mRNAs of a hairpin element responsible for ribosome dependent pyrrolysine insertion into proteins”. 《Biochimie》 87 (9–10): 813–817. doi:10.1016/j.biochi.2005.03.006. PMID 16164991.
- ↑ 가 나 Lu, Y.; Freeland, S. (2006). “On the evolution of the standard amino-acid alphabet”. 《Genome Biology》 7 (1): 102. doi:10.1186/gb-2006-7-1-102. PMC 1431706. PMID 16515719.
- ↑ Voet, D.; Voet, J. G. (2004). 《Biochemistry》 3판. John Wiley & Sons. ISBN 978-0471193500.
- ↑ Chakauya, E.; Coxon, K. M.; Ottenhof, H. H.; Whitney, H. M.; Blundell, T. L.; Abell, C.; Smith, A. G. (2005). “Pantothenate biosynthesis in higher plants”. 《Biochemical Society Transactions》 33 (4): 743–746. doi:10.1042/BST0330743. PMID 16042590.
- ↑ 가 나 다 라 마 Weber, A. L.; Miller, S. L. (1981). “Reasons for the occurrence of the twenty coded protein amino acids”. 《Journal of Molecular Evolution》 17 (5): 273–284. Bibcode:1981JMolE..17..273W. doi:10.1007/BF01795749. PMID 7277510. S2CID 27957755.
- ↑ Koyack, M. J.; Cheng, R. P. (2006). 〈Design and Synthesis of β-Peptides With Biological Activity〉. 《Protein Design》. Methods in Molecular Biology 340. 95–109쪽. doi:10.1385/1-59745-116-9:95. ISBN 978-1-59745-116-1. PMID 16957334.
- ↑ Boniface, A.; Parquet, C.; Arthur, M.; Mengin-Lecreulx, D.; Blanot, D. (2009). “The Elucidation of the Structure of Thermotoga maritima Peptidoglycan Reveals Two Novel Types of Cross-link”. 《Journal of Biological Chemistry》 284 (33): 21856–21862. doi:10.1074/jbc.M109.034363. PMC 2755910. PMID 19542229.
- ↑ Arias, C. A.; Martín-Martínez, M.; Blundell, T. L.; Arthur, M.; Courvalin, P.; Reynolds, P. E. (1999). “Characterization and modelling of vanT: A novel, membrane-bound, serine racemase from vancomycin-resistant Enterococcus gallinarum BM4174”. 《Molecular Microbiology》 31 (6): 1653–1664. doi:10.1046/j.1365-2958.1999.01294.x. PMID 10209740. S2CID 25796469.
- ↑ Gao, X.; Chooi, Y. H.; Ames, B. D.; Wang, P.; Walsh, C. T.; Tang, Y. (2011). “Fungal Indole Alkaloid Biosynthesis: Genetic and Biochemical Investigation of the Tryptoquialanine Pathway in Penicillium aethiopicum”. 《Journal of the American Chemical Society》 133 (8): 2729–2741. doi:10.1021/ja1101085. PMC 3045477. PMID 21299212.
- ↑ Padmanabhan, S.; Baldwin, R. L. (1991). “Straight-chain non-polar amino acids are good helix-formers in water”. 《Journal of Molecular Biology》 219 (2): 135–137. doi:10.1016/0022-2836(91)90553-I. PMID 2038048.
- ↑ Ramadan, S. E.; Razak, A. A.; Ragab, A. M.; El-Meleigy, M. (1989). “Incorporation of tellurium into amino acids and proteins in a tellurium-tolerant fungi”. 《Biological Trace Element Research》 20 (3): 225–232. doi:10.1007/BF02917437. PMID 2484755. S2CID 9439946.
- ↑ Curis, E.; Nicolis, I.; Moinard, C.; Osowska, S.; Zerrouk, N.; Bénazeth, S.; Cynober, L. (2005). “Almost all about citrulline in mammals”. 《Amino Acids》 29 (3): 177–205. doi:10.1007/s00726-005-0235-4. PMID 16082501. S2CID 23877884.
- ↑ Vermeer, C. (1990). “Gamma-carboxyglutamate-containing proteins and the vitamin K-dependent carboxylase”. 《The Biochemical Journal》 266 (3): 625–636. doi:10.1042/bj2660625. PMC 1131186. PMID 2183788.
- ↑ Bhattacharjee, A; Bansal, M (2005). “Collagen structure: The Madras triple helix and the current scenario”. 《IUBMB Life》 57 (3): 161–172. doi:10.1080/15216540500090710. PMID 16036578. S2CID 7211864.
- ↑ Park, M. H. (2006). “The post-translational synthesis of a polyamine-derived amino acid, hypusine, in the eukaryotic translation initiation factor 5A (eIF5A)”. 《Journal of Biochemistry》 139 (2): 161–169. doi:10.1093/jb/mvj034. PMC 2494880. PMID 16452303.
- ↑ Blenis, J; Resh, M. D. (1993). “Subcellular localization specified by protein acylation and phosphorylation”. 《Current Opinion in Cell Biology》 5 (6): 984–989. doi:10.1016/0955-0674(93)90081-z. PMID 8129952.
- ↑ Copley, S. D.; Frank, E.; Kirsch, W. M.; Koch, T. H. (1992). “Detection and possible origins of aminomalonic acid in protein hydrolysates”. 《Analytical Biochemistry》 201 (1): 152–157. doi:10.1016/0003-2697(92)90188-D. PMID 1621954.
- ↑ Van Buskirk, J. J.; Kirsch, W. M.; Kleyer, D. L.; Barkley, R. M.; Koch, T. H. (1984). “Aminomalonic acid: Identification in Escherichia coli and atherosclerotic plaque”. 《Proceedings of the National Academy of Sciences》 81 (3): 722–725. Bibcode:1984PNAS...81..722V. doi:10.1073/pnas.81.3.722. PMC 344907. PMID 6366787.
- ↑ Dasuri, K.; Ebenezer, P. J.; Uranga, R. M.; Gavilán, E.; Zhang, L.; Fernandez-Kim, S. O. K.; Bruce-Keller, A. J.; Keller, J. N. (2011). “Amino acid analog toxicity in primary rat neuronal and astrocyte cultures: Implications for protein misfolding and TDP-43 regulation”. 《Journal of Neuroscience Research》 89 (9): 1471–1477. doi:10.1002/jnr.22677. PMC 3175609. PMID 21608013.
- ↑ Trown, P. W.; Smith, B.; Abraham, E. P. (1963). “Biosynthesis of cephalosporin C from amino acids”. 《The Biochemical Journal》 86 (2): 284–291. doi:10.1042/bj0860284. PMC 1201751. PMID 13994319.