메발론산 경로
메발론산 경로(영어: mevalonate pathway) 또는 아이소프레노이드 경로(영어: isoprenoid pathway) 또는 HMG-CoA 환원효소 경로(영어: HMG-CoA reductase pathway)는 진핵생물, 고균 및 일부 세균에 존재하는 필수적인 대사 경로이다.[1] 메발론산 경로는 아이소펜테닐 피로인산(IPP)과 다이메틸알릴 피로인산(DMAPP)이라고 불리는 2가지 5탄소 대사 중간생성물을 생성하며, 이들 물질들은 아이소프레노이드를 만드는데 사용된다. 아이소프레노이드는 콜레스테롤, 헴, 비타민 K, 조효소 Q10 및 모든 스테로이드 호르몬들과 같은 30,000가지가 넘는 다양한 생체분자들을 총칭한다.[2]

메발론산 경로는 아세틸-CoA에서부터 시작해서 아이소펜테닐 피로인산과 다이메틸알릴 피로인산의 생성으로 끝난다.[3] 콜레스테롤 합성 저해제인 스타틴은 메발론산 경로(메발로네이트 경로)내에서 HMG-CoA 환원효소(HMG-CoA reductase)를 억제한다.
개요 편집
메발론산 경로의 전체적인 개요는 다음의 그림과 같다.
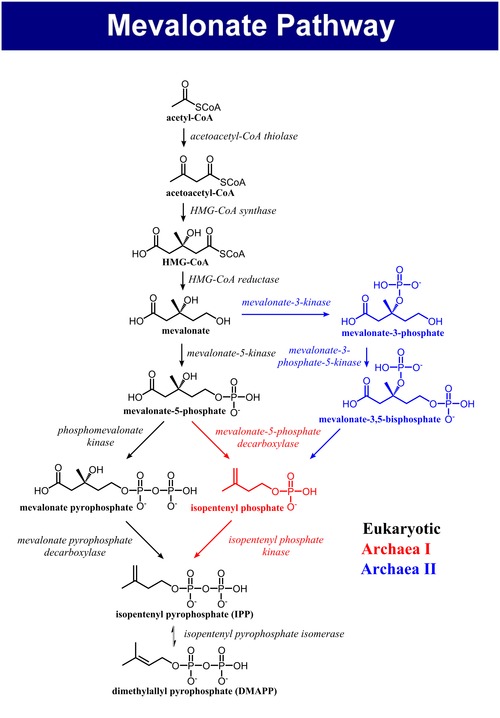 |
아세틸-CoA로부터 메발론산의 생성 편집
메발론산 경로는 진핵생물, 고균, 세균에서 모두 같은 방식으로 시작된다. 메발론산 경로의 유일한 탄소 공급원은 아세틸-CoA이다. 첫 번째 단계는 싸이올레이스에 의해 2분자의 아세틸-CoA를 축합시켜 아세토아세틸-CoA를 생성하는 것이다. 이어서 아세토아세틸-CoA와 아세틸-CoA가 HMG-CoA 생성효소에 의해 축합되어 β-하이드록시 β-메틸글루타릴-CoA(HMG-CoA)를 형성하는 반응이 일어난다. 다음으로 HMG-CoA 환원효소에 의한 HMG-CoA의 환원으로 메발론산을 생성한다.[4]
메발론산으로부터 활성화된 아이소프렌 단위로의 전환 편집
메발론산을 아이소펜테닐 피로인산(IPP)과 다이메틸알릴 피로인산(DMAPP)으로 전환시키는 경로에는 3가지 종류가 있다. 진핵생물에서는 메발론산의 5번 탄소의 하이드록시기에서 두 번 인산화되고 탈카복실화되어 아이소펜테닐 피로인산이 생성된다.[4] 할로페락스 볼카니(Haloferax volcanii)와 같은 일부 고균에서 메발론산은 5번 탄소의 하이드록시기에서 한 번 인산화되고, 탈카복실화되어 아이소펜테닐 인산(IP)을 생성하고, 이 후에 다시 인산화되어 아이소펜테닐 피로인산(IPP)을 생성한다(고균의 메발론산 경로 I).[5] 테르모플라스마 아치도필룸(Thermoplasma acidophilum)에서 발견된 고균의 또 다른 메발론산 경로는 메발론산의 3번 탄소의 하이드록시기에서 인산화된 다음, 5번 탄소의 하이드록시기에서 인산화된다. 이러한 결과로 생성되는 대사산물인 메발론산 3,5-이중인산은 탈카복실화되어 아이소펜테닐 인산(IP)이 된 다음, 인산화되어 아이소펜테닐 피로인산(IPP)을 생성한다(고균의 메발론산 경로 II).[6][7]
조절 및 피드백 편집
몇몇 핵심 효소들은 스테롤 조절요소 결합 단백질(SREBP)의 활성화에 대한 DNA 전사 조절을 통해 활성화될 수 있다. 이러한 세포 내 센서는 낮은 콜레스테롤 수치를 감지하여 메발론산 경로에 의한 내인성 생성을 자극할 뿐만 아니라 LDL 수용체를 상향 조절함으로써 지질단백질의 흡수를 증가시킨다. 또한 메발론산 경로의 조절은 mRNA의 번역 속도, 환원효소의 분해 및 인산화를 조절함으로써 달성된다.[1]
약리학 편집
많은 약물들이 메발론산 경로를 표적으로 삼는다.
질병 편집
메발론산 경로에 영향을 미치는 질환들은 다음과 같다.
- 메발론산 키네이스 결핍증
- 메발론산뇨증
- 면역글로불린 D 증후군 (HIDS)
대체 경로 편집
식물, 대부분의 세균, 말라리아 원충과 같은 일부 원생동물들은 비메발론산 경로라고 불리는 대체 경로를 사용하여 아이소프레노이드들을 생성할 수 있다.[8] 메발론산 경로와 비메발론산 경로의 생성물은 둘 다 아이소펜테닐 피로인산(IPP)과 다이메틸알릴 피로인산(DMAPP)으로 동일하지만, 아세틸-CoA를 아이소펜테닐 피로인산으로 전환시키는 효소 반응은 완전히 다르다. 고등 식물에서 비메발론산 경로는 색소체에 존재하며, 메발론산 경로는 세포질에 존재한다.[8] 비메발론산 경로를 가지고 있는 세균의 예로는 대장균 및 결핵균과 같은 병원균이 있다.
효소 반응 편집
| 효소 | 반응 | 설명 |
|---|---|---|
| 싸이올레이스 |  |
아세틸-CoA는 또 다른 아세틸-CoA 분자와 축합하여 아세토아세틸-CoA를 생성한다. |
| HMG-CoA 생성효소 |  |
아세토아세틸-CoA는 또다른 아세틸-CoA 분자와 축합하여 β-하이드록시 β-메틸글루타릴-CoA(HMG-CoA)를 생성한다. |
| HMG-CoA 환원효소 |  |
HMG-CoA는 NADPH에 의해 메발론산으로 환원된다. 이 과정은 콜레스테롤 합성의 속도 제한 단계이기 때문에, HMG-CoA 환원효소는 의약품(예: 스타틴)의 표적이 된다. |
| 메발론산 키네이스 |  |
메발론산은 5번 탄소의 하이드록시기에서 인산화되어 5-포스포메발론산(메발론산 5-인산이라고도 함)을 생성한다. |
| 메발론산-3-키네이스 |  |
메발론산은 3번 탄소의 하이드록시기에서 인산화되어 메발론산 3-인산을 생성한다. |
| 메발론산-3-인산-5-키네이스 |  |
메발론산 3-인산은 5번 탄소의 하이드록시기에서 인산화되어 메발론산 3,5-이중인산을 생성한다. |
| 포스포메발론산 키네이스 |  |
5-포스포메발론산이 인산화되어 5-피로포스포메발론산을 생성한다. |
| 피로포스포메발론산 탈카복실화효소 |  |
5-피로포스포메발론산은 탈카복실화되어 아이소펜테닐 피로인산(IPP)을 생성한다. |
| 아이소펜테닐 피로인산 이성질화효소 |  |
아이소펜테닐 피로인산(IPP)은 다이메틸알릴 피로인산(DMAPP)으로 이성질화된다. |
같이 보기 편집
각주 편집
- ↑ 가 나 Buhaescu I, Izzedine H (2007) Mevalonate pathway: areview of clinical and therapeutical implications. ClinBiochem 40:575–584.
- ↑ Holstein, S. A., and Hohl, R. J. (2004) Isoprenoids: Remarkable Diversity of Form and Function. Lipids 39, 293−309
- ↑ Goldstein, J. L., and Brown, S. B. (1990) Regulation of the mevalonate pathway. Nature 343, 425−430
- ↑ 가 나 Miziorko H (2011) Enzymes of the mevalonate pathway of isoprenoid biosynthesis. Arch Biochem Biophys 505:131-143.
- ↑ Dellas, N., Thomas, S. T., Manning, G., and Noel, J. P. (2013) Discovery of a metabolic alternative to the classical mevalonate pathway. eLife 2, e00672
- ↑ Vinokur JM, Korman TP, Cao Z, Bowie JU (2014) Evidence of a novel mevalonate pathway in archaea. Biochemistry 53:4161–4168.
- ↑ Azami Y, Hattori A, Nishimura H, Kawaide H, YoshimuraT, Hemmi H (2014) (R)-mevalonate-3-phosphate is an intermediate of the mevalonate pathway in Thermoplasma acidophilum. J Biol Chem 289:15957–15967.
- ↑ 가 나 Banerjee A, Sharkey TD. (2014) Methylerythritol 4-phosphate (MEP) pathway metabolic regulation. Nat Prod Rep 31:10431055
외부 링크 편집
- Rensselaer Polytechnic Institute Archived 2017년 7월 4일 - 웨이백 머신 page on cholesterol synthesis (including regulation)